Hyperglycemia is considered to be a key factor in the development of atherosclerosis and other vascular complications, both considered major causes of morbidity and mortality in diabetic patients. Prolonged exposure to hyperglycemia may promote long-term damage and dysfunction of blood vessels and various cell types, leading to vasculopathy. Hemoglobin A1c (HbA1c), a clinical measure of glycemic control, has been correlated with both macrovascular and microvascular complications in type 2 diabetes. Higher HbA1c is positively correlated with increased rates of vascular complications, especially in the microvasculature.1 Moreover, prolonged hyperglycemia can also accelerate the formation of advanced glycation end products (AGE), formed by spontaneous non-enzymatic reactions between reducing sugars and proteins, DNA or lipids. The level of AGE in the form of methylglyoxal has been demonstrated to increase up to 6- and 3-fold in the serum of type 1 and type 2 diabetic patients, respectively.2 AGE have been recently linked to the incident rate of vascular complications in diabetes.3 Accumulated AGE have the potential to activate multiple vasculopathy-related signaling pathways.4,5 AGE act via their receptors, RAGE, which have been found to be expressed on neutrophil plasma membranes.6 Engagement of AGE and their receptors activate various signal transduction pathways, resulting to increased reactive oxygen species (ROS) production and accelerated atherogenesis in diabetes.7 During the inflammatory response, AGE enhance the expression of adhesion molecules, including vascular cell adhesion molecule-1 (VCAM-1), intracellular adhesion molecule-1 (ICAM-1) and E-selectin, which contribute to increased leukocyte adhesion to endothelial cells.8 AGE-RAGE interactions have been shown to increase extravasation of macromolecules in diabetic rats in vivo, suggesting that AGEs may modulate vascular permeability.9 Moreover, AGEs have been detected in both extra- and intra-cellular localizations of atherosclerotic lesions.10
Neutrophils have been shown to have functional defects in diabetic patients.11 Although the role of neutrophils in the pathogenesis of diabetes and its complications is not yet fully understood, they are now thought to be part of the development and progression of diabetes-related atherosclerosis via several mechanisms. The presence of excess ROS is widely accepted as a substantial mediator in the development of diabetes-related vasculopathy. Neutrophils are one of the major sources of ROS. In response to inflammatory stimuli, activated neutrophils adhere and transmigrate through vascular endothelial wall toward inflammatory sites. Binding of neutrophil surface molecules and their ligands on vascular endothelial cells via selectins and integrins promote rolling and adherence of neutrophils to the endothelium. Integrins are heterodimers composed of alpha and beta subunits. The leukocyte integrin macrophage-1 antigen (MAC-1 integrin, CD11b/CD18 or αmβ2 integrin) plays an important role during transmigration by firmly adhering to ICAM-1 expressed on the endothelial cells. Mice deficient in P-selectin and ICAM-1 have been shown to have decreased atherosclerotic lesion formation.12 Since these molecules are important in neutrophil adhesion to endothelium, the association between these molecules and plaque formation may imply the role of neutrophils in atherogenesis. CD66 was also found to be upregulated in neutrophils during inflammation. Engagement of CD66 with a CD66 monoclonal antibody also increased oxidative burst and β2 integrin-mediated adhesion, indicating that CD66 regulates neutrophil functions.13
Myeloperoxidase (MPO) is an enzyme most abundantly found in azurophilic granules of neutrophils. Upon neutrophil activation, MPO is released and subsequently binds to the CD11b/CD18 integrins on the neutrophil surface. This was found to induce degranulation, CD11b expression and nicotinamide adenine dinucelotide phosphate (NADPH) oxidase activity.14 In addition, MPO-positive neutrophils were observed in the lesional cap and adventitia layer of murine atherosclerotic lesions in the intermediate and advanced stages of atherogenesis.15 The level of MPO secreted by neutrophils in plasma has also been demonstrated to be correlated with coronary artery disease.16 The excess production of cytokines and enzymes and prolonged inflammation may contribute to inappropriate vascular activation and tissue damage.
While there is more to be understood about the role of neutrophils in vasculopathy, the aforementioned evidence indicates that neutrophils may play an important role in the development of diabetes-related atherosclerosis. In the present study, we sought to investigate the effect of hyperglycemia and AGEs on the functions of neutrophils, which may be responsible for the inflammatory state in diabetes and related to diabetes-associated atherosclerosis.
Subjects
The study included 12 type 2 diabetic subjects, aged 35 to 65 years, who were diagnosed and treated at the Diabetes Clinic of the Siriraj Hospital in Thailand. Five healthy subjects for in vitro studies were recruited from the local population and university staff.
The healthy subjects and diabetic patients were evaluated and subsequently excluded if with any of the following: history of clinically significant peripheral arterial disease, cardiovascular disease or cerebrovascular disease; abnormal lipid profile; overweight; obesity; taking immunosuppressive or immune-stimulating drug; pregnant; or lactating. In accordance with the level of HbA1c, diabetic patients were divided into 2 groups: those with 7% or lower HbA1c were designated as good control (n=6) and more than 9.0% HbA1c as poor control (n=6). Subject characteristics are shown in Table 1.
Click here to download Figure 1Table 1. Metabolic parameters by subject group.
All subjects signed a written informed consent after being informed of the objectives and procedures of the study. The study protocol was approved by the Siriraj Institutional Review Board.
Isolation of neutrophilsVenous blood samples collected from subjects were placed in tubes containing ethylenediaminetetraacetic acid (EDTA) for the isolation and cultivation of neutrophils. Neutrophils were isolated by Ficoll-Hypaque density gradient separation and dextran sedimentation. Briefly, 30 mL of venous blood was mixed with a double volume of Dulbecco's phosphate-buffered saline (Gibco® DPBS, Invitrogen Corporation, California, USA); 30 mL of this diluted sample was layered over 10 mL of Histopaque®-1077 (Sigma-Aldrich Co. LLC, Missouri, USA) in a 50 mL conical plastic centrifuge tube. After centrifugation at 800 x g for 30 minutes at room temperature, the plasma and mononuclear cell-rich interphase was removed. The pellet containing polymorphonuclear leukocytes (PMN) and red blood cells was mixed with an equal volume of 3% dextran in 0.9% sodium chloride solution and allowed to stand for one hour at room temperature. PMN-rich supernatant was collected then centrifuged for 7 minutes at 1500 rpm at 4°C. The resulting supernatant was discarded. Contaminating erythrocytes were removed by hypotonic lysis using sterile water. The neutrophils were harvested and resuspended in DPBS. Purity of neutrophils was determined by differential staining with Giemsa stain. The cell viability was more than 95% as assessed by trypan blue exclusion.
Treatment of neutrophilsNeutrophils from healthy individuals were cultured in Roswell Park Memorial Institute 1640 Medium (Gibco® RPMI 1640 Medium, Invitrogen Corporation, California, USA) supplemented with 10% fetal bovine serum (Gibco® FBS), 2 mM L-glutamine (Gibco® L-glutamine), 100 μg/mL penicillin and 100 IU/mL streptomycin (Sigma-Aldrich Co. LLC, Missouri, USA). Cells were incubated under different conditions at 37°C in a 5% CO2 atmosphere for 1 hour: 100 μg/mL bovine serum albumin (Calbiochem® BSA, EMD, Darmstadt, Germany), 100 μg/mL advanced glycation end product-BSA (Calbiochem® AGE-BSA, Merck Millipore, New Jersey, USA), 5 mM D-glucose (Sigma-Aldrich Co. LLC) and 25 mM D-glucose. For cell activation, cells were incubated with 100 ng/mL phorbol 12-myristate 13-acetate (PMA, Sigma-Aldrich Co. LLC, Missouri, USA) at 37°C for 15 minutes for dihydrorhodamine (DHR) assay, or 1 hour for other experiments.
Determination of reactive oxygen speciesROS production was determined by DHR assay within one hour after isolation.17 Approximately 1 x 106 cells/mL of isolated PMNs were mixed with DHR (Invitrogen Corporation, California, USA) and stimulated with PMA for 15 minutes. For control cells, Hank's balanced salt solution (Gibco® HBSS, BRL, California, USA) was added instead of PMA and incubated at 37°C in a shaking water bath for 15 minutes.
The ability to produce ROS, determined by the level of DHR conversion to rhodamine, was measured by BD FACSort™ flow cytometer (Becton Dickinson Immunocytometry Systems, California, USA). At least 30,000 cells of neutrophils were collected. Neutrophils were gated by their characteristic forward and side scatter profiles, which represent size and granularity, respectively. Data analysis was performed by using Cell Quest Software (BD Biosciences, California, USA).
Flow cytometry for CD11b and CD66b expressionCell surface molecules were identified using BD FACSCalibur™ flow cytometer (Becton Dickinson Immunocytometry Systems, California, USA). For each analysis, approximately 1 x 106 cells were stained with fluorescein isothiocyanate (FITC)-conjugated mouse anti-CD66b or phycoerythrin (PE)-conjugated mouse anti-CD11b monoclonal antibodies (BD Biosciences, California, USA). Mouse isotype-matched Ig was used as negative control.
Neutrophils were initially gated by forward scatter (FSC) and side scatter (SSC) for determining positive expression intensity of FITC- and PE-conjuated antibodies. Data were collected for 50,000 cells/sample and analyzed by Cell Quest Software (BD Biosciences, California, USA).
Detection of myeloperoxidaseNeutrophils from all subjects were incubated with PMA or left untreated for one hour. The level of MPO enzyme in culture supernatants of neutrophils was quantified by commercially available high-sensitivity enzyme-linked immunosorbent assay (ELISA) kit (Assay Designs, Inc., Michigan, USA). The assays were performed as described in the manufacturer's protocol.
Cell migration assayCell migration was evaluated by the induction of cell migration towards interleukin-8 through Corning® Transwell® polycarbonate membrane (6.5 mm diameter and 5.0 μm pore polycarbonate membrane insert) (Corning Inc., New York, USA). Approximately 1 x 106 neutrophils were incubated in the upper chambers of the wells filled with 200 μL of serum-free RPMI-1640 medium under different conditions: 5 mM glucose, 25 mM glucose, 100 μg/mL BSA and AGE-BSA in the presence of PMA. The lower chambers of the wells were filled with 100 ng/mL of IL-8 in 600 μL of serum-free RPMI-1640 medium. Cells were allowed to migrate through the membrane to the lower chambers for one hour at 37°C in a 5% CO2 atmosphere. Migrated cells in the 600 μL suspension were harvested from the lower chambers, washed and resuspended in 500 μL fixing solution. These were then counted by flow cytometry gating only PMN populations according to the FSC and SSC profiles.
Neutrophil-endothelial cell adhesion assayNeutrophil-endothelial cell adhesion was evaluated by the quantification of adherent neutrophils to an endothelial cell monolayer. The endothelial hybrid cell line EA.hy 926 was seeded at a concentration of 1 x 105 cells/well into 24-well plates. These were incubated in Dulbecco's Modified Eagle Medium: Nutrient Mixture F-12 (Gibco® DMEM-F12K medium, Invitrogen Corporation, California, USA) supplemented with 10% FBS (Gibco® FBS), 100 μg/mL penicillin and 100 IU/mL streptomycin (Sigma-Aldrich Co. LLC, Missouri, USA) for 24 hours at 37°C until the cells were 80% confluent.
Neutrophils from healthy subjects were isolated and incubated in different conditions: 5 mM glucose, 25 mM glucose, 100 μg/mL BSA or 100 μg/mL AGE-BSA at 37°c for one hour. After incubation, 1 x 106 cells/mL of neutrophils were then added to each well of the endothelial monolayer and allowed to adhere for 30 minutes at 37°C. Non-adhered neutrophils were removed by gentle wash twice with warmed DPBS. After washing, cells were fixed in 1% paraformaldehyde. The adherence of neutrophils to endothelial cell monolayer was quantified by counting in 5 randomly selected high-power fields (x400) per well under a phase contrast microscope.
Statistical analysisValues were normally distributed and data were reported as mean ± SEM, unless otherwise stated. Comparisons between the subject groups were done by Mann-Whitney Test using SPSS PASW Statistics 18. Differences between groups in the in vitro experiments were evaluated by student's t test. A p-value less than or equal to 0.05 was considered statistically significant.
Clinical parameters
The average level of HbA1c was 7.1 ± 0.5% in the group of diabetic patients with good glycemic control and 10.4 ± 0.9% in those with poor glycemic control (PCD). Other than HbA1c, there were no statistically significant differences between the 2 groups of diabetic patients for the rest of the metabolic parameters, which were within normal limits. These findings suggest that hyperglycemia was the only factor for atherosclerosis in the diabetic patients in the study.
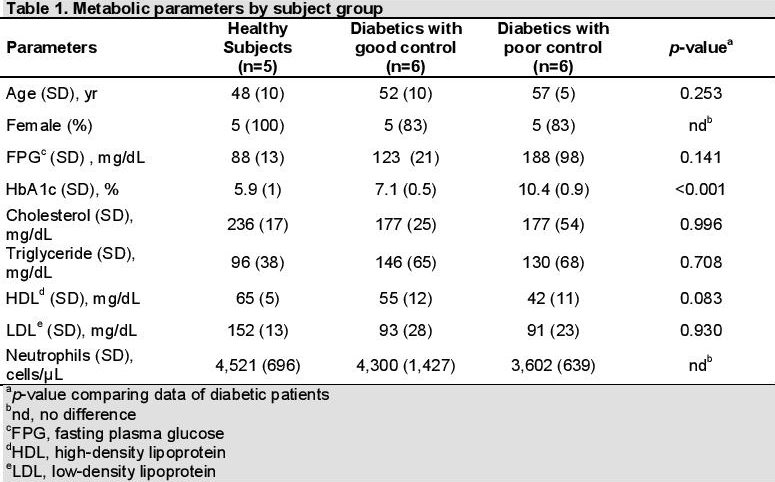
Neutrophil ROS productionPhysiologic production of ROS is important in the degradation of phagocytosed materials, cell signaling and homeostasis. Overproduction of ROS, known as oxidative stress, results to significant damage to other molecules and cell structures. We investigated the effect of glycemic control in diabetic patients on neutrophil oxidative burst function. ROS production by neutrophils was assessed by DHR assay. Interestingly, the production of basal and PMA-stimulated ROS in neutrophils from diabetics with poor control was significantly lower compared to those with good control (p-value=0.014) (Figure 1a).
Isolated neutrophils from healthy subjects were exposed to high glucose concentration (25 mM) or AGE-BSA compared to physiologically normal glucose concentration (5 mM) or unmodified BSA to assess the effect of high glucose and AGEs, simulating the diabetic milieu. Basal and PMA-stimulated ROS production were measured. We observed that basal and PMA-stimulated ROS production of neutrophils incubated in high glucose or AGE-BSA were not different from those incubated in normal glucose and BSA (Figures 1b and 1c).
F Click here to download Figure 1Figure 1. Neutrophil oxidative burst function among healthy subjects, diabetics with good control and diabetics with poor control by dihydrorhodamine (DHR) assay. A significantly lower basal and stimulated reactive oxygen species (ROS) production was seen in diabetics with poor control (a) (*p=0.05). Isolated neutrophils from healthy subjects incubated in physiologic (5 mM) and high glucose (25 mM) concentrations (b) and in bovine serum albumin (BSA) and advanced glycation end product-BSA (AGE-BSA) (c), showing similar basal and stimulated ROS production..
Neutrophil expression of CD11b and CD66b
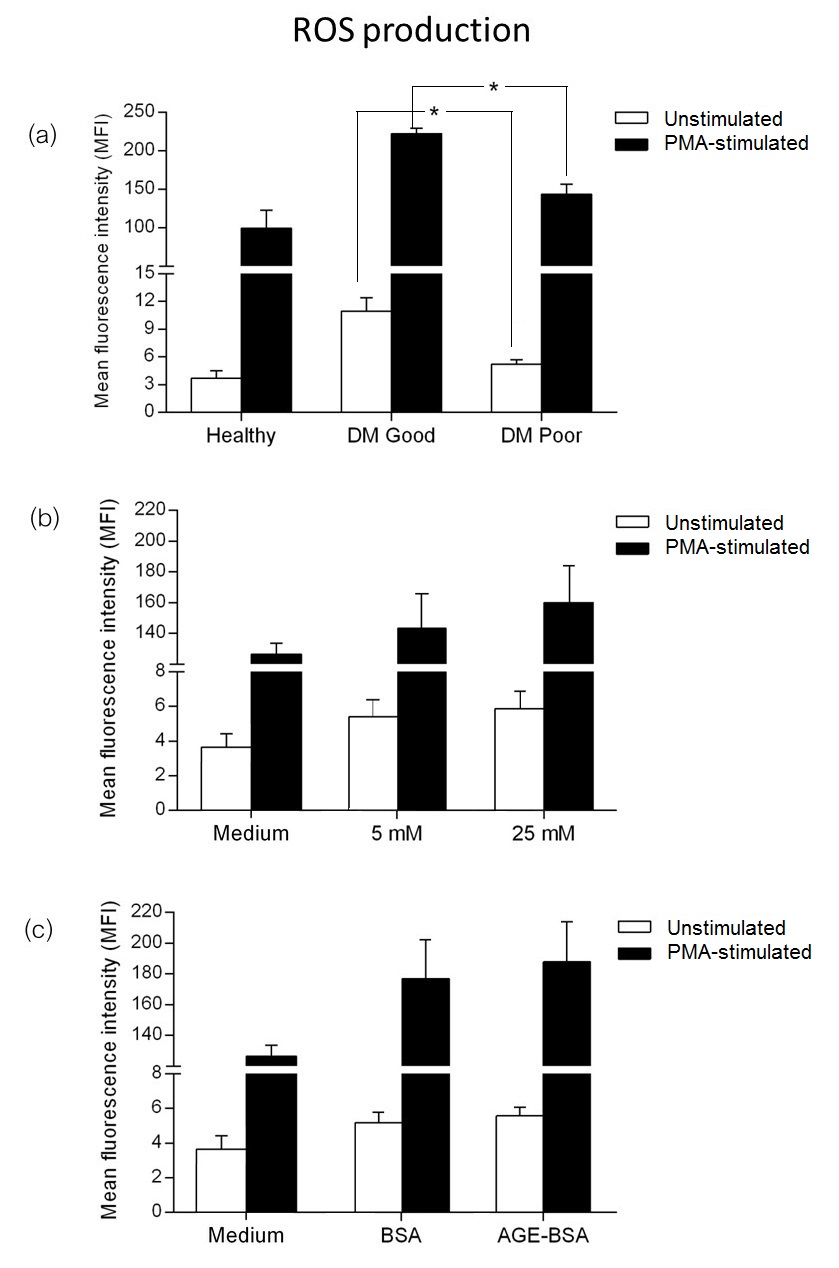
During inflammation, neutrophils upregulate surface molecule expression in response to stimuli. We measured the expression of activation markers CD11b and CD66b, known to be involved in vascular inflammation, on neutrophils from diabetic patients with different levels of glycemic control.
Basal expression of CD11b was lower in diabetics compared to healthy subjects. Among diabetics, neutrophils from those with poor glycemic control had significantly higher CD11b expression compared to those with good control (p-value=0.021) (Figure 2a). Upon PMA stimulation, neutrophils from both groups of diabetics were found to have a markedly increased expression of CD11b, with a significantly higher expression in diabetics with poor control (p-value=0.034). Similarly, basal expression of CD66b in diabetics was lower than in healthy subjects. CD66b expression in diabetics with poor glycemic control was significantly higher compared to those with good control(Figure 2b).
Isolated neutrophils from healthy subjects were incubated in physiologic (5 mM) and high (25 mM) concentrations of glucose. After short-term incubation in vitro, CD11b and CD66b expression were comparable in both glucose conditions (Figures 2c and 2d). Similarly, basal and PMA-stimulated expressions of CD11b and CD66b were also similar in AGE-BSA and unmodified BSA (Figure 2e and 2f).
Click here to download Figure 2Figure 2.Expression of activation markers. Surface molecules CD11b and CD66b in neutrophils from healthy subjects, diabetics with good control and diabetics with poor control (a and b), showing lower basal expression of CD11b and CD66b in diabetics, and increased expression after phorbol 12-myristate 13-acetate (PMA) stimulation (*p≤0.05). Isolated neutrophils from healthy subjects incubated in physiologic (5 mM) and high (25 mM) glucose concentrations (c and d) and in bovine serum albumin (BSA) and advanced glycation end product-BSA (AGE-BSA) (e and f)
Neutrophil migration
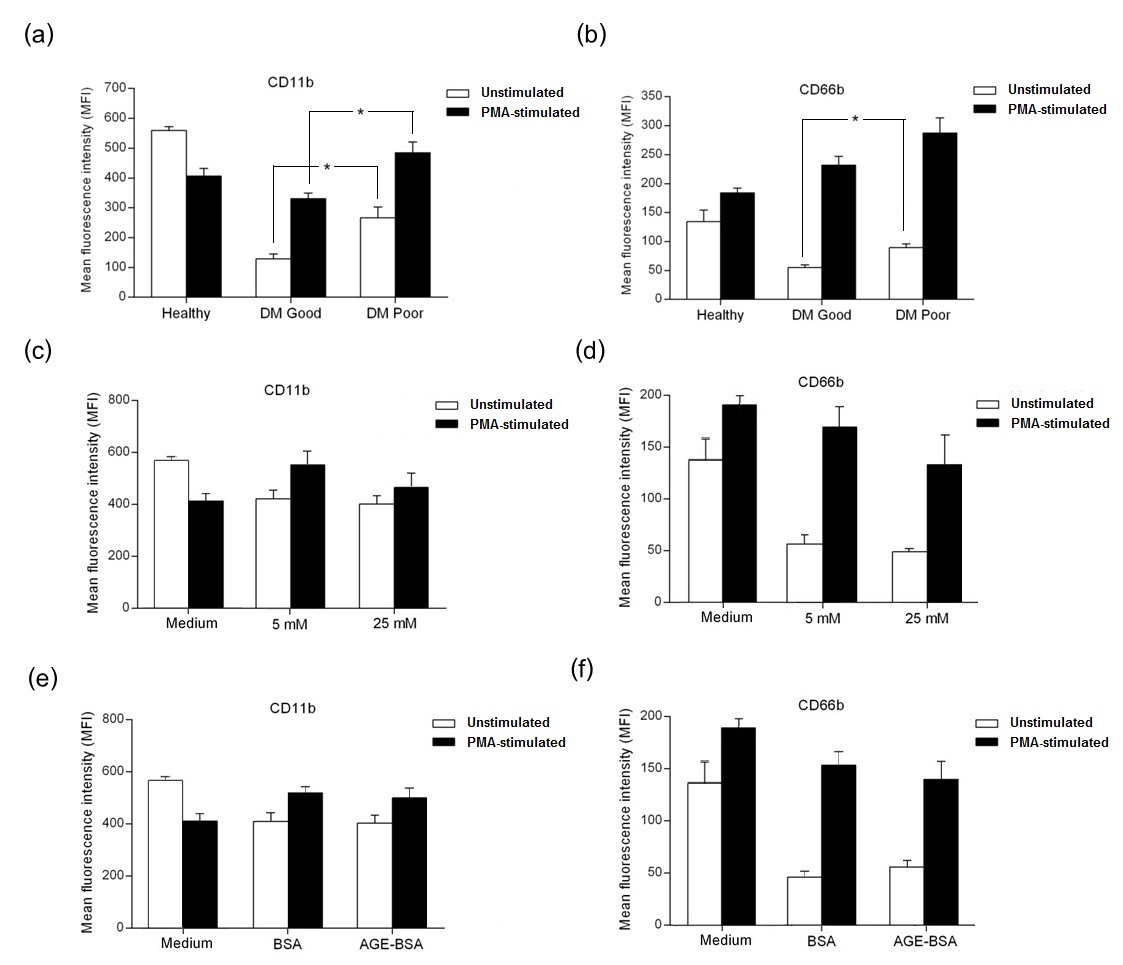
The effect of high glucose and AGE-BSA on IL-8-induced chemotaxis of neutrophils from healthy subjects was also observed. Cell migration was performed by the induction of neutrophil migration through Transwell® membrane towards IL-8. Neutrophils were incubated in 5 mM and 25 mM glucose in the presence of PMA for 1 hour. Migrated cells in lower chamber were counted by flow cytometry. The migration assay showed a significant increase in neutrophil migration in high glucose conditions. A slightly higher number of neutrophils migrating towards IL-8 was seen in the presence of AGE-BSA compared to unmodified BSA (Figure 3).
Click here to download Figure 3Figure 3.IL-8-induced migration of neutrophils, showing increased chemotaxis in high glucose conditions (*p=0.05).
Neutrophil-endothelial cell adhesion
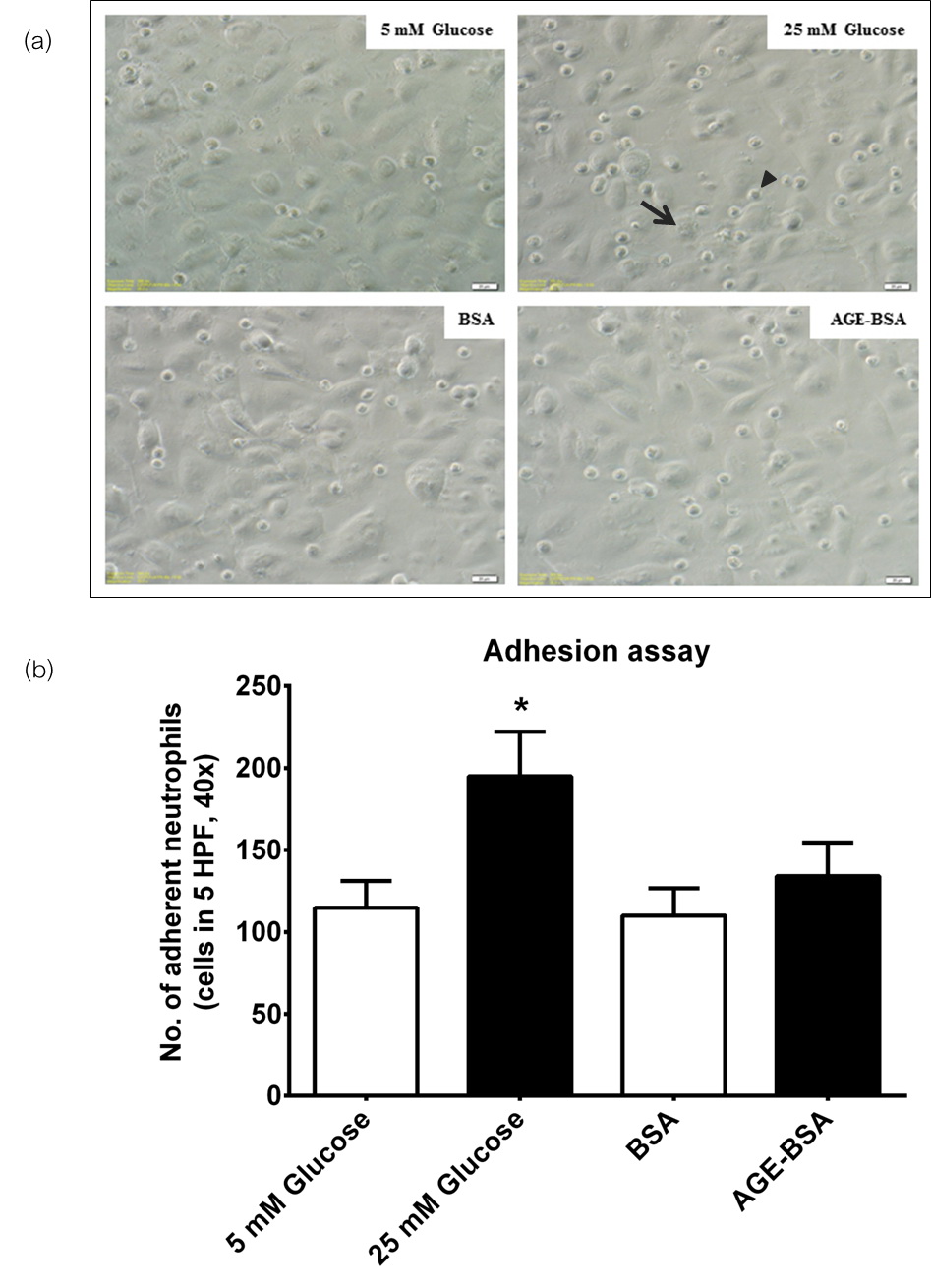
Incubation of neutrophils from healthy subjects in high glucose concentrations significantly increased adherence to EA.hy926 endothelial cells (Figure 4). There was a slight increase in the number of adherent cells exposed to AGE-BSA compared to unmodified BSA, but this did not reach statistical significance. These observations may suggest that the increase in neutrophil adherence to endothelial cells may promote the initial interaction between these cells under hyperglycemic conditions.
Click here to download Figure 4Figure 4.Neutrophil-endothelial adhesion assay. a, photomicrograph of EA.hy926 monolayer (arrow) under phase contrast microscopy showing adherent neutrophils (arrow head) (x400). b, comparison of adherent neutrophils from healthy subjects, showing increased adherence under high glucose conditions (*p≤0.05).
Myeloperoxidase release from activated neutrophils
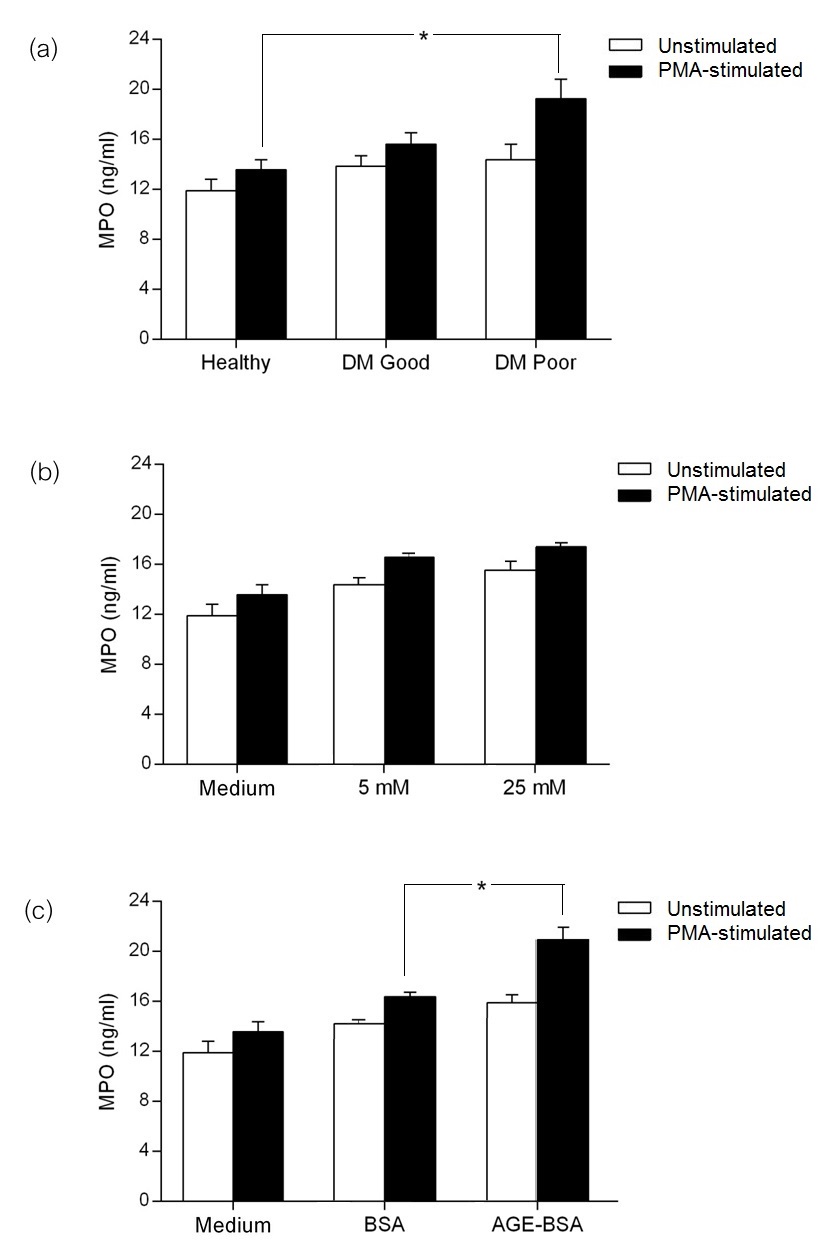
Basal and PMA-stimulated amounts of MPO in culture supernatant were measured by high-sensitivity sandwich ELISA. The basal level of MPO production was similar across healthy subjects, diabetics with good control and diabetics with poor control (Figure 5a). Upon PMA stimulation, MPO production was significantly higher in the supernatant with neutrophils from diabetics with poor control compared to cells from healthy subjects. In in vitro studies, MPO production of neutrophils from healthy subjects incubated in 25 mM glucose was comparable to that of cells incubated in 5 mM glucose concentration (Figure 5b). In the presence of AGE-BSA, PMA-stimulated neutrophils had significantly higher MPO release compared to unmodified BSA (Figure 5c).
Click here to download Figure 5Figure 5.Myeloperoxidase (MPO) production of neutrophils in healthy subjects, diabetics with good control and diabetics with poor control. Highest levels of MPO were seen in diabetics with poor control after phorbol 12-myristate 13-acetate (PMA) stimulation (a). Isolated neutrophils from healthy subjects incubated in physiologic (5 mM) and high (25 mM) glucose concentrations (b), showing comparable levels of MPO production in both conditions. Incubation in advanced glycation end product-bovine serum albumin (AGE-BSA) was shown to have caused a significantly higher production of MPO after PMA stimulation (c) (*p≤0.05).
In this study, we have demonstrated the effect of hyperglycemia on the functions of neutrophils, both ex vivo from patients and in vitro. We evaluated the activation and functional status of neutrophils, which may be related to the pro-inflammatory state in diabetic patients and the subsequent development of atherosclerosis. In our ex vivo studies of neutrophils from diabetic subjects, cells from those with good control exhibited significantly higher ROS production compared to those with poor control. Neutrophils from diabetics with poor glycemic control showed significantly increased expression of activation markers CD11b and CD66b, as well as higher MPO production upon PMA stimulation.
On the other hand, in our in vitro studies on neutrophils from healthy subjects, the release of MPO was significantly increased after PMA stimulation in those cells incubated in AGE-BSA, compared to unmodified BSA. We have demonstrated for the first time that incubation of neutrophils from healthy subjects in a high glucose environment significantly enhanced neutrophil migration and adherence to endothelial cells.
Upon activation by inflammatory stimuli, neutrophils produce increased ROS. Enhanced production of ROS causes oxidative stress, which is thought to lead to the development of the diabetic vascular complications. We found higher ROS production in diabetic patients with good glycemic control compared to healthy subjects. This was consistent with findings in another study demonstrating that neutrophils had enhanced production of ROS in type 2 diabetes.18 Interestingly, both basal and PMA-stimulated ROS production from diabetics with poor glycemic control were significantly lower than in diabetics with good control. As there has never been any evidence on the effect of glycemic variability on ROS production before, we have only one possible explanation why diabetics with poor control had reduced neutrophil ROS production compared tho those with good control in this study. Inhibition of GAPDH activity activates the protein kinase C (PKC) pathway of hyperglycemic damage in endothelial cells.19 This can also lead to activation of the ROS production pathway. In our unpublished data (ongoing manuscript preparation), sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) of neutrophil homogenates from diabetics with poor glycemic control expressed higher glyceraldehyde 3-phophate dehydrogenase (GAPDH) than those with good control. This may explain the reduced production of ROS in diabetics with poor glycemic control with higher GAPDH. However, ROS production in poorly controlled diabetics was higher compared to healthy subjects. To determine which component of hyperglycemia has an effect on the cells, neutrophils from healthy subjects were incubated in high glucose (25 mM) or AGE-BSA in vitro. Similar levels of ROS generation were observed after short term exposure of neutrophils to high glucose and AGE-BSA, compared to ROS production after incubation in normal glucose (5 mM) and unmodified BSA. This may be due to a short period of incubation, since ROS production has been shown to significantly increase with AGE concentrations greater than 200 μg/mL and incubation time up to 4 hours in a recently published study.20
Basal expression of CD11b and CD66b in both groups of diabetic patients was lower in comparison to healthy subjects. A possible explanation is that prolonged hyperglycemia may induce a resting state in the degradation of surface molecules. The results were in accordance with our in vitro findings, which showed a slightly decreased expression of surface markers in a high glucose environment (25 mM). To support this hypothesis, we incubated cells up to 8 hours and found that CD11b and CD66b were not further upregulated upon stimulation with PMA, compared to 1-hour incubation. A previous study also demonstrated that PMA stimulation in vitro resulted in a significantly decreased expression of CD11b in a time-dependent manner, suggesting that prolonged stimulation may lead to loss of surface molecules by proteolytic degradation.21 Interestingly, expression of CD11b and CD66b from PCD was significantly upregulated compared to those with good control. After PMA, increased expression of CD11b and CD66b was observed in neutrophils from both diabetic groups, but this was significantly higher in cells from diabetics with poor control. In the in vitro experiments, neutrophils from healthy subjects were incubated in RPMI-1640 medium, which has a glucose concentration of 11 mM. This may explain why there were no significant differences between cells incubated in 5 mM and 25 mM glucose, as well as AGE-BSA versus BSA. Our results were consistent with other previous studies. Increased expression of CD11b and CD66b molecules in neutrophils has been reported in diabetic patients.22,23 CD66b expression in neutrophils from type 2 diabetics was significantly upregulated compared to healthy subjects. However, some studies have shown controversial data for CD11b expression. In patients with diabetes, CD11b expression in neutrophils was higher at resting state but no difference was found after activation with fMLP.24 Basal expression of CD11b in neutrophils showed no difference in type 2 diabetic subjects compared to normal subjects, but the expression was elevated in diabetic subjects upon activation with PMA.25 A previous study demonstrated that the expression of CD11b in diabetic and normal subjects was not different, but was significantly lower in diabetic subjects after PMA activation.6
The surface molecule CD11b is used for firm adhesion with endothelial cells during transmigration, while CD66b is believed to interact with extracellular matrix, regulating neutrophil function and mediating the release of IL-8, a cytokine essential for the recruitment and accumulation of PMNs.8,26 Elevated CD11b and CD66b expression in poorly controlled patients may promote neutrophil migration and adhesion to endothelial cells. We investigated the ability of neutrophils to migrate towards IL-8 and to adhere onto cells of an endothelial monolayer. We were able to demonstrate that high glucose significantly enhanced cell migration and adherence to endothelial cells. This is in agreement with another study that found high glucose enhanced endothelial cell capacity for neutrophil adherence by increasing endothelial cell adhesion molecules.27
We measured the ability of neutrophils to release myeloperoxidase. MPO is a peroxidase enzyme found most abundant in azurophilic granules of neutrophils. Several studies have shown the relationship between MPO and inflammatory processes in vascular disease. MPO was seen in atherosclerotic lesions.28 MPO has also been investigated as a biomarker for inflammation in acute coronary syndromes.29 In our study, we observed significantly higher MPO production in neutrophils from poorly controlled diabetics compared to healthy subjects. Other studies have reported that plasma levels of MPO in type 2 diabetes were increased compared to subjects without diabetes.30,31 In accordance with our ex vivo findings, our in vitro study showed significantly higher PMA-activated MPO production under exposure to AGE-BSA. The results suggest that AGEs formed in diabetics with poor glycemic control may be a potential factor for the stimulation of MPO production. The link between the altered neutrophil functions and the pathogenesis of diabetes-related atherosclerosis is still being investigated.
We have observed activated neutrophil function, particularly ROS and MPO production in diabetic patients, and increased adhesive capacity by way of CD11b and CD66b neutrophil expression in hyperglycemic conditions in vitro. Our study was limited by our small sample size and the possibility of other unidentified confounding factors that may influence the observed alterations in immunity and neutrophil function. At this time we cannot conclude from the findings data of the in vitro study whether the alterations seen in neutrophil function are due to conditions simulating short-term hyperglycemia (incubation in high glucose concentration), or an environment akin to chronic hyperglycemia (incubation in AGE-BSA). Nevertheless, we were able to demonstrate for the first time that incubation of neutrophils in a high glucose environment enhanced cell migration towards IL-8 and adherence to endothelial cells. These altered functions of neutrophils may, at least in part, be involved in the development and progression of atherosclerosis in diabetic patients.
The authors declare that they have no competing interest that may lend bias to this study.
AcknowledgementsThis study was supported by the Office of the Higher Education Commission and Mahidol University under the National Research Universities Initiative, and The Thailand Research Fund (RTA5380005). W. Umsa-ard was partially supported by the Siriraj Graduate Thesis Scholarships. J. Soongsathitanon was the recipient of the Siriraj Chalermprakiat Fund. The EA.hy926 cell line was a gift kindly received from Dr. Sa-nga Pattanakitsakul. The authors wish to thank Dr. Egarit Noulsri and Miss Surada Lerdwana for technical assistance in flow cytometry.
1. Stratton IM, Adler AI, Neil HA, et al. Association of glycaemia with macrovascular and microvascular complications of type 2 diabetes (UKPDS 35): Prospective observational study. BMJ. 2000;321(7258):405-12.http://dx.doi.org/ 10.1136/bmj.321.7258.405.
2. McLellan AC, Thornalley PJ, Benn J, et al. Glyoxalase system in clinical diabetes mellitus and correlation with diabetic complications. Clin Sci (Lond). 1994;87(1):21-9.http://dx.doi.org/10.1042/cs0870021.
3. Nin JW, Jorsal A, Ferreira I, et al. Higher plasma levels of advanced glycation end products are associated with incident cardiovascular disease and all-cause mortality in type 1 diabetes: A 12-year follow-up study. Diabetes Care. 2011;34(2):442-7.http://dx.doi.org/10.2337/dc10-1087.
4. Wada R, Yagihashi S. Role of advanced glycation end products and their receptors in development of diabetic neuropathy. Ann N Y Acad Sci. 2005;1043:598-604.
5. Al-Mesallamy HO, Hammad LN, El-Mamoun TA, et al. Role of advanced glycation end product receptors in the pathogenesis of diabetic retinopathy. J Diabetes Complications. 2011;25(3):168-74.
6. Collison KS, Parhar RS, Saleh SS, et al. RAGE-mediated neutrophil dysfunction is evoked by advanced glycation end products (AGEs). J Leukoc Biol. 2002;71(3):433-44.
7. Mullarkey CJ, Edelstein D, Brownlee M. Free radical generation by early glycation products: A mechanism for accelerated atherogenesis in diabetes. Biochem Biophys. Res Commun 1990;173(3):932-9.
8. Basta G, Lazzerini G, Massaro M, et al. Advanced glycation end products activate endothelium through signal-transduction receptor RAGE: A mechanism for amplification of inflammatory responses. Circulation. 2002;105(7):816-22.http://dx.doi.org/ 10.1161/hc0702.104183.
9. Bonnardel-Phu E, Wautier JL, Schmidt AM, et al. Acute modulation of albumin microvascular leakage by advanced glycation end products in microcirculation of diabetic rats in vivo. Diabetes.1999;48(10):2052-8.http://dx.doi.org/ 10.2337/diabetes.48.10.2052.
10. Horiuchi S, Sano H, Higashi T, et al. Extra- and intracellular localization of advanced glycation end-products in human atherosclerotic lesions. Nephrol Dial Transplant. 1996;11(Suppl 5):81-6.http://dx.doi.org/ 10.1093/ndt/11.supp5.81.
11. Delamaire M, Maugendre D, Moreno M, et al. Impaired leucocyte functions in diabetic patients. Diabet Med. 1997;14(1):29-34.http://dx.doi.org/10.1002/(SICI)1096-9136(199701)14:1<29::AID-DIA300>3.0.CO;2-V.
12. Collins RG, Velji R, Guevara NV, et al. P-Selectin or intercellular adhesion molecule (ICAM)-1 deficiency substantially protects against atherosclerosis in apolipoprotein E-deficient mice. J Exp Med. 2000;191(1):189-94.http://dx.doi.org/ 10.1084/jem.191.1.189.
13. Ruchaud-Sparagano MH, Stocks SC, Turley H, et al. Activation of neutrophil function via CD66: differential effects upon beta 2 integrin mediated adhesion. Br J Haematol. 1997;98(3):612-20.http:// 10.1046/j.1365-2141.1997.2523070.x.
14. Lau D, Mollnau H, Eiserich JP, et al. Myeloperoxidase mediates neutrophil activation by association with CD11b/CD18 integrins. Proc Natl Acad Sci USA. 2005;102(2):431-6.http://dx.doi.org/ 10.1073/pnas.0405193102.
15. van Leeuwen M, Gijbels MJ, Duijvestijn A, et al. Accumulation of myeloperoxidase-positive neutrophils in atherosclerotic lesions in LDLR-/- mice. Arterioscler Thromb Vasc Biol. 2008;28(1):84-9.http://dx.doi.org/ 10.1161/ATVBAHA.107.154807.
16. Zhang R, Brennan ML, Fu X, et al. Association between myeloperoxidase levels and risk of coronary artery disease. JAMA. 2001;286(17):2136-42.
17. Vowells SJ, Sekhsaria S, Malech HL, et al. Flow cytometric analysis of the granulocyte respiratory burst: a comparison study of fluorescent probes. J Immunol Methods. 1995;178(1):89-97.
18. Hand WL, Hand DL, Vasquez Y. Increased polymorphonuclear leukocyte respiratory burst function in type 2 diabetes. Diabetes Res Clin Pract. 2007;76(1):44-50.
19. Du X, Matsumura T, Edelstein D, et al. Inhibition of GAPDH activity by poly (ADP-ribose) polymerase activates three major pathways of hyperglycemic damage in endothelial cells. J Clin Invest. 2003;112(7):1049-57.http://dx.doi.org/ 10.1172/JCI18127.
20. Bansal S, Siddarth M, Chawla D, et al. Advanced glycation end products enhance reactive oxygen and nitrogen species generation in neutrophils in vitro. Mol Cell Biochem. 2012;361(1-2):289-96.
21. Davey PC, Zuzel M, Kamiguti AS, et al. Activation-dependent proteolytic degradation of polymorphonuclear cells. Br J Haematol. 2000;111:934-42.http://dx.doi.org/ 10.1111/j.1365-2141.2000.02401.x.
22. Grykiel K, Zozulinska D, Kostrzewa A, et al. [Evaluation of expression of polymorphonuclear neutrophil surface receptors in patients with type 1 diabetes]. Pol Arch Med Wewn. 2001;105:377-81.
23. van Oostrom AJ, van Wijk JP, Sijmonsma TP, et al. Increased expression of activation markers on monocytes and neutrophils in type 2 diabetes. Neth J Med. 2004;62(9):320-5.
24. Rao KM, Hatchell DL, Cohen HJ, et al. Alterations in stimulus-induced integrin expression in peripheral blood neutrophils of patients with diabetic retinopathy. Am J Med Sci. 1997;313:131-7.
25. Senior PA, Marshall SM, Thomas TH. Dysregulation of PMN antigen expression in Type 2 diabetes may reflect a generalized defect of exocytosis: Influence of hypertension and microalbuminuria. J Leukoc Biol. 1999;65(6):800-7.
26. Schröder AK, Uciechowski P, Fleischer D, et al. Crosslinking of CD66B on peripheral blood neutrophils mediates the release of interleukin-8 from intracellular storage. Hum Immunol. 2006;67(9):676-82.
27. Omi H, Okayama N, Shimizu M, et al. Participation of high glucose concentrations in neutrophil adhesion and surface expression of adhesion molecules on cultured human endothelial cells: Effect of antidiabetic medicines. J Diabetes Complications. 2002;16(3):201-8.http://dx.doi.org/10.1016/S1056-8727(01)00163-5.
28. Daugherty A, Dunn JL, Rateri DL, et al. Myeloperoxidase, a catalyst for lipoprotein oxidation, is expressed in human atherosclerotic lesions. J Clin Invest. 1994;94(1):437-44.http://dx.doi.org/ 10.1172/JCI117342.
29. Loria V, Dato I, Graziani F, et al. Myeloperoxidase: A new biomarker of inflammation in ischemic heart disease and acute coronary syndromes. Mediators Inflamm. 2008; 2008:135625.
30. Hatanaka E, Monteagudo PT, Marrocos MS, et al. Neutrophils and monocytes as potentially important sources of proinflammatory cytokines in diabetes. Clin Exp Immunol. 2006;146:443-7.http://dx.doi.org/ 10.1111/j.1365-2249.2006.03229.x.
31. Wiersma JJ, Meuwese MC, van Miert JN, et al. Diabetes mellitus type 2 is associated with higher levels of myeloperoxidase. Med Sci Monit. 2008;14(8):CR406-10.
Authors are required to accomplish, sign and submit scanned copies of the JAFES Declaration that the article represents original material that is not being considered for publication or has not been published or accepted for publication elsewhere.
Consent forms, as appropriate, have been secured for the publication of information about patients; otherwise, authors declared that all means have been exhausted for securing such consent.
The authors have signed disclosures that there are no financial or other relationships that might lead to a conflict of interest. All authors are required to submit Authorship Certifications that the manuscript has been read and approved by all authors, and that the requirements for authorship have been met by each author.